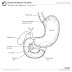
Eyes in various animals show adaptation to their requirements. For example, birds of prey have much greater visual acuity than humans, and some can see ultraviolet light. The different forms of eye in, for example, vertebrates and molluscs are often cited as examples of parallel evolution, despite their distant common ancestry.
The very earliest "eyes", called eyespots, were simple patches of photoreceptor protein in unicellular animals. In multicellular beings, multicellular eyespots evolved, physically similar to the receptor patches for taste and smell. These eyespots could only sense ambient brightness: they could distinguish light and dark, but not the direction of the light source.
Through gradual change, as the eyespot depressed into a shallow "cup" shape, the ability to slightly discriminate directional brightness was achieved by using the angle at which the light hit certain cells to identify the source. The pit deepened over time, the opening diminished in size, and the number of photoreceptor cells increased, forming an effective pinhole camera that was capable of dimly distinguishing shapes.
The thin overgrowth of transparent cells over the eye's aperture, originally formed to prevent damage to the eyespot, allowed the segregated contents of the eye chamber to specialise into a transparent humour that optimised colour filtering, blocked harmful radiation, improved the eye's refractive index, and allowed functionality outside of water. The transparent protective cells eventually split into two layers, with circulatory fluid in between that allowed wider viewing angles and greater imaging resolution, and the thickness of the transparent layer gradually increased, in most species with the transparent crystallin protein.
The gap between tissue layers naturally formed a bioconvex shape, an optimally ideal structure for a normal refractive index. Independently, a transparent layer and a nontransparent layer split forward from the lens: the cornea and iris. Separation of the forward layer again formed a humour, the aqueous humour. This increased refractive power and again eased circulatory problems. Formation of a nontransparent ring allowed more blood vessels, more circulation, and larger eye sizes.